―しがない農学徒の雑記帳―
しがない農学徒が日々思うところを書き散らします。項目
色に対する反応のモデル化
はじめに
「飛んで火にいる夏の虫」とは、自ら災難に向かう者をきらめく炎に近づき焼け死ぬ虫に例えたことわざのことです。このことわざのように、昆虫は走光性という、光に対して近づく性質をもちます。走光性の中でも、昆虫種によっては光の波長組成に対する選好性、すなわち波長選好性がみられます。例えば、多くのハエ目昆虫は紫外域を好みます。この性質を利用して、紫外蛍光管による電撃殺虫機が衛生害虫や不快害虫の駆除の為に用いられています。
一方、農学においては波長選好性を用いて農業害虫や天敵の行動を制御し、作物を保護するという「光防除」が近年提唱されています(霜田,2014)。この背景の1つとして、発光ダイオード (LED) が比較的安価に入手できるようになり、光環境を操作する事が現実的になったということが挙げられます。
光防除の実用化を念頭に、多くの昆虫種で波長選好性が調べられています(図1)。選好される波長と昆虫の複眼の感度の関係をみてみると、多くの昆虫種は感度が高い波長を選好する事がわかります。一方で、波長選好性の背後にある、昆虫の神経系における情報処理機構に言及した研究例は多くありません。研究で得たデータを用いて、様々な光源のもとにおける昆虫の行動を予測するためには、その情報処理機構をモデル化するという研究手法があります。ここでは、その方法を紹介します。
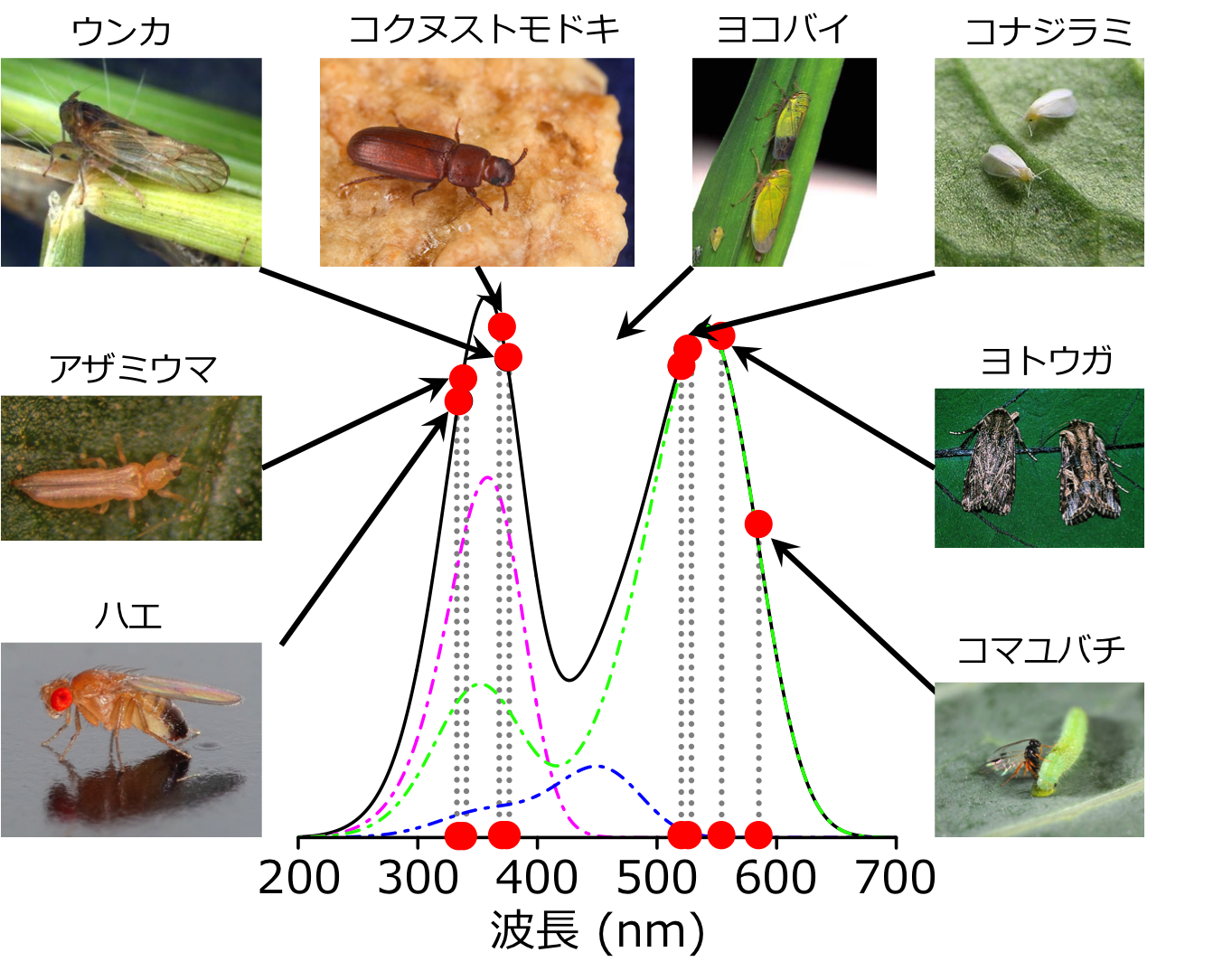 図1 複眼の感度スペクトルと波長選好性の関係。紫外、青、緑といった感度型の視細胞の感度スペクトル(破線)の重ね合わせで複眼の感度スペクトル(実線)が決まります。複眼の感度スペクトルは紫外域(550 nm 付近)か緑域(350 nm 付近)、またはその両方に極大をもつが、そのどちらかの波長を多くの昆虫種は選好します。この図では、仮想的な感度スペクトルに対して複数の昆虫種の選好波長を示しています。
図1 複眼の感度スペクトルと波長選好性の関係。紫外、青、緑といった感度型の視細胞の感度スペクトル(破線)の重ね合わせで複眼の感度スペクトル(実線)が決まります。複眼の感度スペクトルは紫外域(550 nm 付近)か緑域(350 nm 付近)、またはその両方に極大をもつが、そのどちらかの波長を多くの昆虫種は選好します。この図では、仮想的な感度スペクトルに対して複数の昆虫種の選好波長を示しています。光受容と波長認識のモデル化
光の波長組成は、動物には色として知覚されます。本節では、光源から放射される光が眼において受容され、脳で知覚されるまで(図2a)を順に説明します。
光源からは放射される光は、プリズムに通す事で単波長の光に分けられます(分光)。光源によって分光後の光の組成に違いがあり、例えばキセノンランプには満遍なく波長成分が含まれます。一方、発光ダイオード(LED)では特定の波長しか含まれない事が多いです。こうした、光が波長λの成分をどれだけ含むかは、放射スペクトルといわれ、L(λ) と表記されます(図2b)。これは、分光測定機で測定できます。
昆虫の眼は複眼であり、複眼の網膜には視細胞が含まれます。視細胞にはいくつかの型があり、型によって吸収しやすい波長や、吸収しにくい波長があります。多くの昆虫は紫外光、青光、そして緑光を吸収する型の視細胞をもちます。ここで、光の波長λの成分をどれだけ吸収するかは、感度スペクトルといわれ、 S(λ) と表記されます(図2b)。これは、電気生理実験により測定できます(蟻川ら、2014)。
放射スペクトルが L(λ) である光が、感度スペクトルが S(λ) である視細胞に入射したときに、視細胞が波長全体として吸収する光量 Q (Q≥0)は、これらの積の波長積分、 Q= ∫L(λ)S(λ) dλ として計算できます。それぞれの型の視細胞で、受容した光量 Q に応じた信号が脳に伝えられます(図2b)。
視細胞からの信号は、脳に含まれる神経回路で処理され、色の認識に至ります。人間では、光の3原色のもとである青、緑、赤の視細胞からの信号に対して、脳では赤と緑、どちらの信号が強いか、あるいは黄(=緑+赤)と青、どちらの信号が強いかが比較されます(池田,1996)。この赤−緑、黄−青という、2つの軸をもとに色が認識されます。この比較は、脳では興奮性の神経と抑制性の神経のはたらきによって行われています。数式では赤−緑、あるいは(緑+赤)−青というように、加算・減算で表現できます。ちなみに、赤と緑の関係や黄と青の関係を反対色と呼びます。昆虫では、ハチ目やハエ目でも、視細胞の型は異なるものの同様のメカニズムが発見されており(Backhaus , 1987; Troje, 1993)、視細胞からの信号に対する加算や減算は、色覚で重要な役割を果たすと考えられています。
この加算・減算を考慮し、波長に対する選好性や波長を識別する能力がモデル化されています(Kelber, 1999; Vorobyev et al. 1998)。
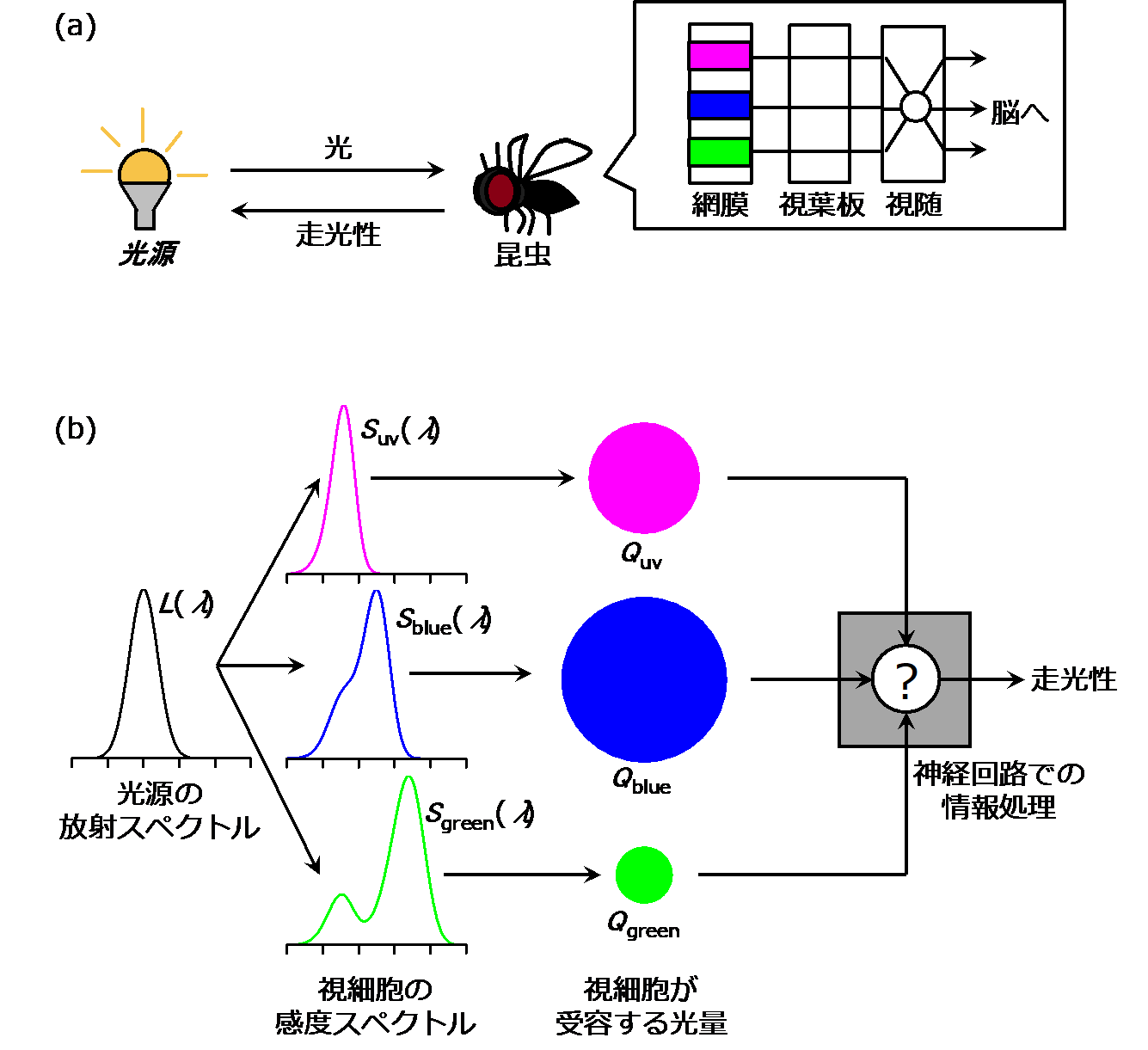 図 2 (a) 光源から放射された光に対して、昆虫は走光性を示します。光は網膜にある様々な感度型の視細胞により受容され、視葉板や視随を通じて脳へ信号が送られます。(b) 光源の放射スペクトルと視細胞の感度スペクトルの積から、視細胞が受容する光量を推測することができます。この光量に応じた信号が神経回路で処理され、走光性が起こります。
図 2 (a) 光源から放射された光に対して、昆虫は走光性を示します。光は網膜にある様々な感度型の視細胞により受容され、視葉板や視随を通じて脳へ信号が送られます。(b) 光源の放射スペクトルと視細胞の感度スペクトルの積から、視細胞が受容する光量を推測することができます。この光量に応じた信号が神経回路で処理され、走光性が起こります。天敵昆虫ブランコヤドリバエにおける波長選好性のモデル化
いくつかの昆虫種で波長選好性はモデル化されていますが、その中でも、ブランコヤドリバエにおける実験・解析(Tokushima et al. 2016)を例として紹介します。交尾を済ませた本種の雌バエは、多くの作物の害虫であるチョウ目の幼虫に対して卵を産みつけます。孵化したウジはこれに寄生し、喰い殺します。この捕食寄生性という生活史があるので、本種は天敵農薬としての利用が期待されています。
まず、本種の選好する波長を調べるため、荻野ら(2015)と同様の実験を行いました。すなわち、紫外、紫、青、緑、橙および赤の LED をプラスチックケースに等間隔に配置し、そこにハエを放した後の行動を観察しました。その結果、本種は他の波長に比べて紫光を顕著に選好する事を発見しました。
この波長選好性がどのようなメカニズムによって定まるか推測する為、視細胞がLED より吸収する光量と情報処理をモデル化しました。ハエ目昆虫の場合は一般に紫外型、すみれ型、青型、緑型、そして広帯域型の視細胞を有します。これらの視細胞が、それぞれの LED よりどれだけの光量を吸収するかを先ほど紹介した積分式で評価しました(図3)。そして、それぞれの視細胞からの信号に対して、仮にどのような「加算・減算」が行われたならば、紫光が選好されるのかをシミュレーションにより推測しました。
シミュレーションに際し、Kelber (1999) に基づき、統計学における一般化線形モデル (Dobson, 2008) の枠組みを用いて、波長選好性を次のようにモデル化しました: C = 1/[1+exp(-η)]
ここで、C は実験における、LED に対する選択率 (0≤C≤1) です。η は、「選好度」のような量です。この式はロジスティック関数といわれ、選好度が大きくなるにつれて選択率も高くなる事を意味します。選好度 η は、視細胞 i (i = 紫外型、すみれ型、青型、緑型または広帯域型)が吸収する光量 Q_i を用いて、次のような1次式でモデル化しました:η=Σb_iQ_i
ここで、b_i (-∞
5 つの型の視細胞のうち、選択率に対して寄与するのはどれか、そして寄与の仕方はどのようであるかを調べるために、全ての視細胞の組み合わせ (2^5=32 通り) につき係数のフィッティングを行いました。この結果、全ての視細胞が波長選好性に寄与し、符号は広帯域型およびすみれ型視細胞がプラス、紫外型、青型および緑型の視細胞がマイナスでした。波長選好性に対して係数がプラスの視細胞は促進的に寄与し、マイナスの視細胞は抑制的に寄与する事で、本種の紫光に対するユニークな選好性がみられたと考えられます。
最も驚くべき事は、広帯域型の視細胞がなければ波長選好性を説明できなかった事です。実は従来、紫外型、紫型、青型および緑型の視細胞は特定波長のみに感度がある為、色覚に関係すると考えられ、一方、後者は広い波長にわたって感度を有する為、形や動きの認識に関係すると考えられていました。しかし、近年ショウジョウバエにおいても広帯域型の視細胞が波長の識別に重要である事が見いだされており、これに次いでブランコヤドリバエでも同じ結論が導きだされました。もしかしたら、広帯域型の視細胞を用いて波長を識別するメカニズムは、進化の過程でハエ目昆虫に共通して備わったのかもしれません。
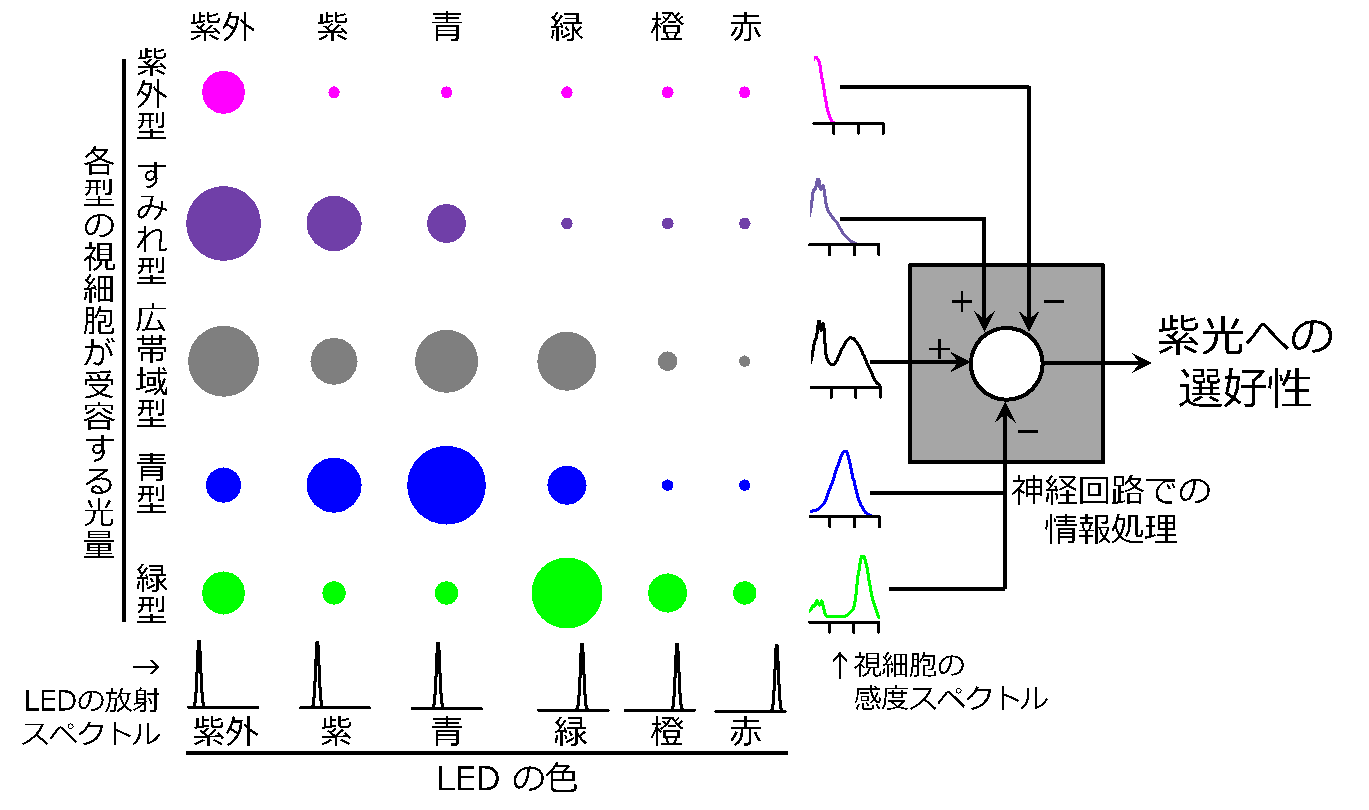 図 3 視細胞(紫外型、すみれ型、広帯域型、青型、緑型)がLED(紫外、紫、青、緑、橙、赤)より受容する光量を円の大きさで示しました。視細胞の感度スペクトルは円の右側に、LED の放射スペクトルは円の下側に示してあります。シミュレーションの結果、走光性に対して広帯域型視細胞とすみれ型視細胞が促進的に、紫外型、青型、緑型視細胞が抑制的に寄与する事で、ブランコヤドリバエは紫光に対して選好すると考えられました。
図 3 視細胞(紫外型、すみれ型、広帯域型、青型、緑型)がLED(紫外、紫、青、緑、橙、赤)より受容する光量を円の大きさで示しました。視細胞の感度スペクトルは円の右側に、LED の放射スペクトルは円の下側に示してあります。シミュレーションの結果、走光性に対して広帯域型視細胞とすみれ型視細胞が促進的に、紫外型、青型、緑型視細胞が抑制的に寄与する事で、ブランコヤドリバエは紫光に対して選好すると考えられました。おわりに
本ページでは、昆虫の光受容や波長選好性のモデル化を説明しました。この手法により、ブランコヤドリバエにおける紫光に対する選好性をつかさどる、意外なメカニズムが発見されました。他の昆虫種に対して同様の実験や解析を行う事で、天敵は誘引するが害虫は寄せ付けないという、光防除に最適な光源を設計できると思われます。